



人胚肾细胞293T
发表时间:2025-07-24人胚肾细胞293T
一. 细胞起源
293T细胞源于人胚胎肾细胞,经腺病毒5(Ad5)DNA片段转化获得,并稳定表达SV40大T抗原(SV40 large T antigen),使其支持含有SV40复制起点的质粒高效复制[1]。
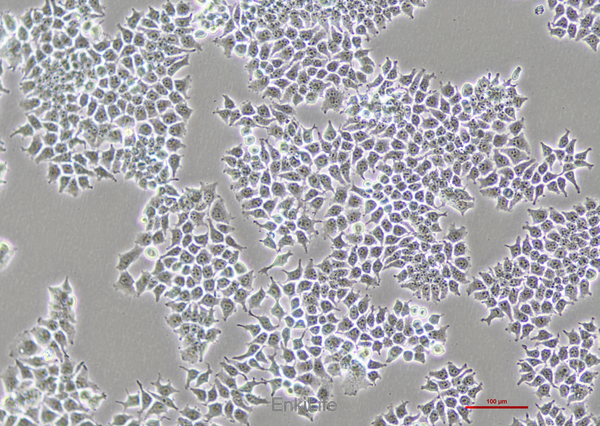
二. 生物学特性
1. 形态与生长特性
-
形态:贴壁生长,呈上皮样或成纤维样,核质比高[2]。
-
增殖能力:分裂速度快(倍增时间约24–36小时),适合大规模培养[1]。
2. 遗传与分子特性
-
SV40大T抗原:增强外源基因表达效率,促进质粒复制[1]。
-
干细胞样表型:在无血清三维球体培养中,高表达ALDH1、CD44+/CD24-标志物,并上调β-catenin、Notch1等干细胞信号通路[3]。
-
间充质转化倾向:三维培养时表达波形蛋白(vimentin)、Zeb1等间充质基因,可能增强转移潜能[3]。
3. 代谢与毒性响应
-
代谢活性:常用于AMPK信号研究,如女贞苷通过提升AMP/ATP比值激活AMPK通路[4]。
-
低毒性耐受:对某些化合物(如共轭亚油酸)无毒性,但对壳聚糖纳米粒等基因载体敏感(抑制率26.08%)[5][6]。
三. 培养与储存
1. 培养基:DMEM+10% FBS+1% P/S[2]。
2. 传代与冻存:常规胰酶消化传代;30%基础培养基+60%FBS+10%DMSO,液氮保存[2]。
3. 转染兼容性:脂质体转染效率高(如GFP载体转染成功率50–60%)[7][8]。
四. 研究应用领域
1. 基因功能与调控研究
-
RNA干扰(RNAi):广泛用于基因沉默(如p21WAF1/CIP1、PES1、HPIP基因),验证靶基因功能[9][10][11]。
-
启动子活性分析:如尼罗罗非鱼β-actin启动子驱动EGFP表达[8]。
-
蛋白互作研究:通过免疫共沉淀筛选互作蛋白(如C14orf166与RS8/EFCB/NRAP)[12]。
2. 疾病机制模型
-
肿瘤研究:模拟癌干细胞表型,用于放疗抗性、转移机制探索[3]。
-
代谢疾病:研究AMPK-脂联素通路(女贞苷激活AMPK)[4]。
-
病毒感染:构建禽流感HA-GFP融合蛋白,分析信号肽对表达的影响[7]。
3. 药物筛选与载体开发
-
抗肿瘤药物:评估化合物选择性(如共轭亚油酸抑制EC109癌细胞)[5]。
-
基因载体:壳聚糖纳米粒转染pGFP,但存在细胞毒性[6]。
-
慢病毒包装:HPIP siRNA慢病毒抑制HeLa/HepG2增殖[10]。
五. 近年研究进展
1. 自噬调控:弓形虫ROP17蛋白在293T中表达,促进血清饥饿诱导的自噬(LC3-II↑、P62↓)[13]。
2. 神经退行性疾病:帕金森研究中用于验证Parkin蛋白调控小胶质细胞NLRP3炎症体[14]。
3. 新型基因工具:构建双标签载体(如pcDNA3.1-Flag-His)用于蛋白互作组学研究[12]。
六. 局限性与克服方法
1. 局限性
-
遗传不稳定性:长期传代可能累积突变,影响实验结果可重复性。
-
非完全正常细胞:表达SV40抗原及间充质基因,不完全代表原代肾细胞[3]。
-
转染毒性:壳聚糖纳米粒等载体导致细胞损伤(抑制率>26%)[6]。
2. 克服方法
-
低代次使用:限制传代次数(<50代),定期鉴定细胞身份。
-
替代载体:优化脂质体或病毒载体(如慢病毒)降低毒性[10]。
-
三维培养模型:利用球体培养模拟体内微环境,提升生理相关性[3]。
七. 总结与展望
293T细胞因其高转染效率、快速增殖及SV40大T抗原特性,已成为分子生物学和疾病研究的核心工具。近年研究拓展至自噬、神经退行性疾病及新型基因编辑领域。未来方向包括:
1. 精准基因编辑:结合CRISPR-Cas9构建更复杂疾病模型。
2. 类器官整合:联合3D培养技术模拟器官微环境。
3. 毒性机制优化:开发低毒载体(如肽纳米粒)提升转染安全性。

参考文献
1.Viebahn, S.Die Rolle der Oberfl?chensialylierung bei der Modulation von Immunantworten.2008.
2.人胚肾细胞293T说明书.武汉恩玑生命科技有限公司.
3.Characterizing cancer cells with cancer stem cell-like features in 293T cells. Debeb BG, et al. Cancer Res. 2010.[PMID: 20663901]
4.郭文文,等.女贞苷通过激活AMPK促进脂联素的组装.2018.
5.吴演,等.海蓬子籽油亚油酸共轭化产物的抗肿瘤作用.2012.
6.王红梅,等.壳聚糖纳米粒基因载体的制备和体外转染实验研究.2010.
7.苏艳,等.信号肽序列对禽流感病毒H5N1 HA蛋白GFP融合分子表达的影响.20010.
8.邹芝英,等.尼罗罗非鱼β-actin基因启动子的分离及其在真核细胞中的活性验证.2011.
9.王朝云,等.利用RNA干扰抑制p21WAF1/CIP1基因的表达及周期阻滞效应[J].生物技术通讯,2008.
10.徐小洁,等.慢病毒介导RNA干扰HPIP蛋白抑制肿瘤细胞增殖.2011.
11.李杰萍,等.利用不同方法检测RNAi抑制人pescadillo基因的表达[J].细胞与分子免疫学杂志,2007.
12.张登禄,等.胰腺癌转移相关基因C14orf166的真核表达及其蛋白相互作用的蛋白质组学筛选[J].中国医药生物技术,2010.
13.刘宏延,等.弓形虫ROP17真核表达载体的构建及ROP17参与自噬的研究.2012.
14.Parkin regulates microglial NLRP3 and represses neurodegeneration in Parkinson's disease. Yan YQ, et al. Research.2025.