



小小凝聚体,掌控突触通讯:SNAP25 LLPS 机制文献解读
发表时间:2026-06-29文献信息
|
|
标题 |
SNAP25 undergoes phase separation to facilitate the assembly of the synaptic vesicle fusion machinery |
|
期刊 |
Cell Reports |
|
|
分区 |
中科院一区 |
|
|
关键词 |
SNAP25, phase separation, SNARE, exocytosis, condensate |
一、研究背景
突触前精准、快速的神经递质释放高度依赖 SNARE 复合物介导的突触胞吐过程,Syntaxin-1、SNAP25、VAMP2 三种核心 SNARE 蛋白相互结合形成四螺旋反式 SNARE 复合体,拉近囊泡与细胞膜距离并启动膜融合进程,SNAP25 结构异常、基因突变会直接破坏突触信号传导,临床中大量发育性癫痫性脑病(DEE)患者体内均检出 SNAP25 杂合突变。既往显微成像研究已经观测到 SNAP25 与 Syntaxin-1 会在突触膜形成纳米尺度蛋白簇,这种聚集状态直接影响囊拴缚、锚定与启动释放,但长期没有研究解释蛋白成簇的分子驱动力;液液相分离(LLPS)作为细胞内无膜区室的组装方式,已被证实调控突触支架蛋白、突触结合蛋白 1 的空间聚集,而作为膜融合核心的 SNARE 蛋白是否依靠相分离实现局部富集始终是领域空白。Min Zhu、Yinghui Liu、Haijia Yu 团队于 2026 年在《Cell Reports》发表题为SNAP25 undergoes phase separation to facilitate the synaptic vesicle fusion machinery的研究,该文结合 SNAP25 独特分子结构 —— 两段 SNARE 基序之间存在大片内在无序连接区,且连接区半胱氨酸可发生棕榈酰化修饰,结合前人 Syntaxin-1 可增强 SNAP25 棕榈酰化的实验结论,提出完整科学假设:SNAP25 依靠无序连接区发生液液相分离,棕榈酰化修饰、Syntaxin-1 共同调控凝聚物形成,相分离产生的液滴可招募全部 SNARE 组分,促进三元复合体组装,DEE 致病突变会破坏相分离并阻断膜融合通路,以此完整填补 SNARE 蛋白聚集机制的研究缺口。
二、研究方法
该研究整合体外生化重构、活细胞光学检测、原代神经元模型、人工膜系统多重实验体系,实验材料涵盖原核表达纯化的野生型与系列突变 SNAP2、Syntaxin-1 胞内段、VAMP2 胞质结构域蛋白,同时构建多种荧光标记融合蛋白、光诱导相分离 OptoDroplet 质粒、SNAP25 与棕榈酰转移酶 ZDHHC17 靶向慢病毒干扰载体;体外实验搭建标准化相分离反应体系,利用荧光共聚焦、DIC 微分干涉成像观察液滴形态,借助沉降实验、浊度定量、圆二色谱、双溴甲烷色氨酸淬灭表征蛋白相分离与构象变化;细胞层面采用 HeLa 细胞敲低回补体系排除过表达干扰,依托 FRAP 荧光漂白恢复实验验证凝聚物液态动态特性,OptoDroplet 光控系统直观观测活细胞内蛋白共凝聚行为;体外膜重构部分构建平面支撑脂双层 SLB、巨型单层囊泡 GUV、小型囊泡 SUV 三种人工膜,模拟细胞膜与突触囊泡结构,还原膜表面蛋白聚集与囊泡招募过程;细胞功能实验使用大鼠原代海马神经元,通过免疫印迹验证蛋白表达量,共聚焦成像观察神经元内蛋白点状聚集;疾病机制部分构建 I67N、I192N 两种 DEE 相关突变蛋白,在 293T 细胞检测 SDS 耐受型功能性 SNARE 复合体生成量,所有实验均设置三次及以上生物学重复,采用 t 检验、单因素方差分析完成统计学差异判定。
三、结果分析
1. 体外与活细胞双重证实 SNAP25 具备液液相分离特性
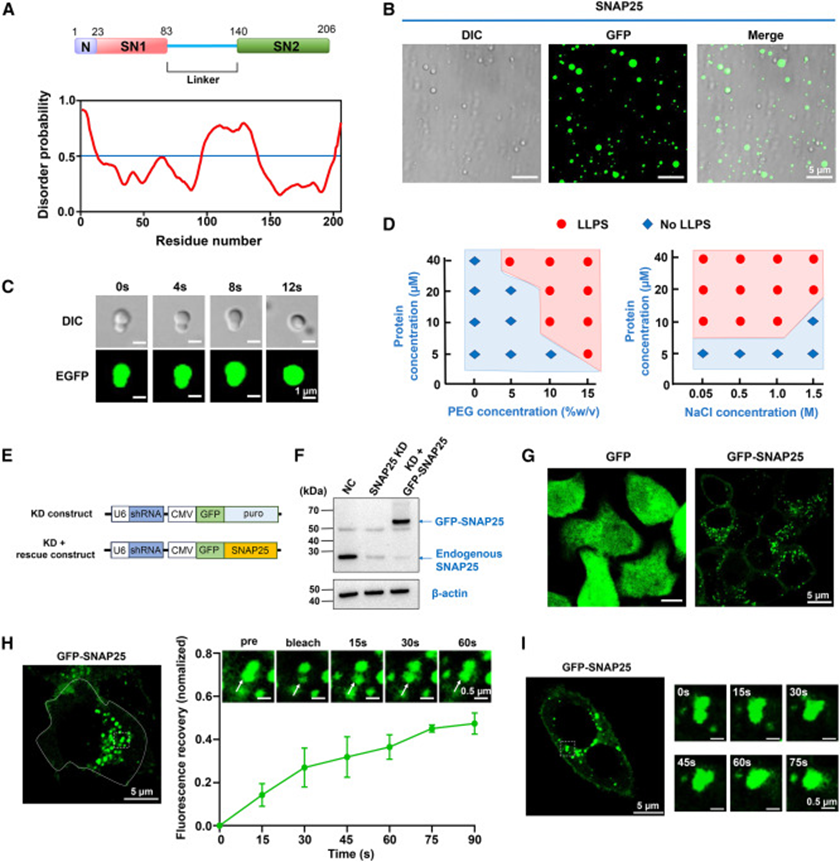
研究首先通过蛋白无序预测工具 PrDOS 解析 SNAP25 氨基酸序列,整条蛋白中两段 SNARE 基序中间的连接区段存在大范围内在无序序列,这也是相分离蛋白的典型结构特征。体外重构体系中,纯化 EGFP 标记 SNAP25 在拥挤剂 PEG 生理模拟环境下自发形成球形液滴,延时成像捕捉到小液滴相互融合的动态画面,直观体现液态属性;梯度浓度、盐离子梯度实验绘制完整相图,证明蛋白浓度、渗透压、分子拥挤程度共同调控相分离发生,生理盐水浓度区间恰好满足凝聚物形成条件。为排除蛋白过表达带来的假聚集,研究构建 SNAP25 敲低 + 内源水平回补 HeLa 细胞,对照组仅表达游离 GFP 时细胞质荧光均匀弥散,而 GFP-SNAP25 细胞内出现大量明亮点状凝聚物;FRAP 漂白实验中凝聚物荧光可缓慢恢复,90 秒达到半数恢复水平,活细胞延时拍摄还能观察点状凝聚物互相融合,从细胞层面印证 SNAP25 在生理表达水平下仍能发生液相凝聚,内外两套体系相互佐证 SNAP25 固有相分离能力。
2. 无序连接区是 SNAP25 相分离的核心必需元件
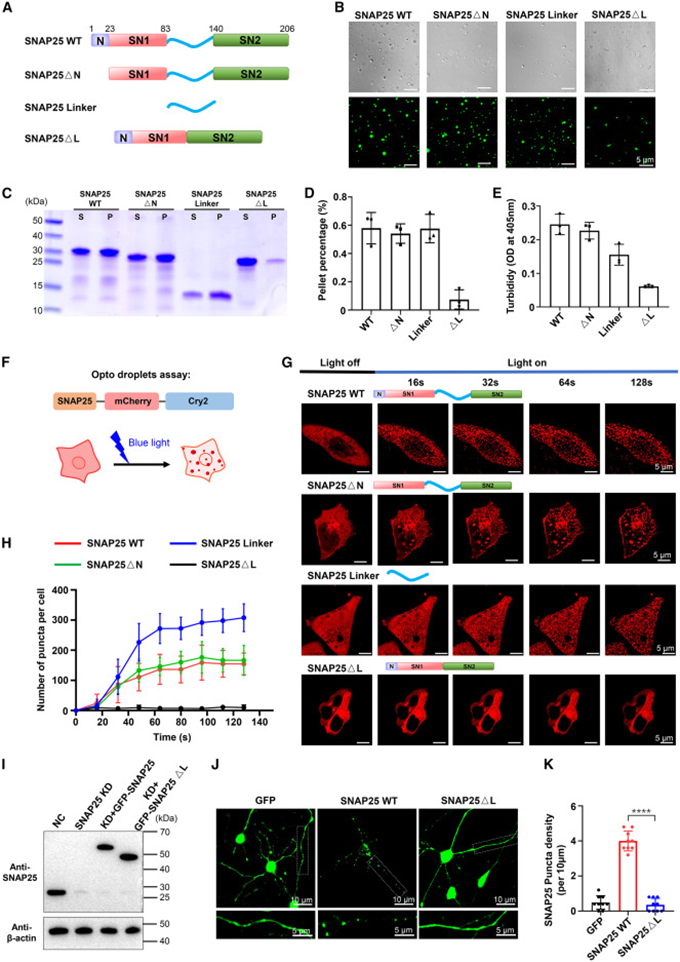
研究者设计系列截短突变体拆解 SNAP25 功能区段,分别去除 N 端短肽、完整删除中间连接区、单独提取连接片段开展平行体外相分离实验。去除 N 端片段的蛋白仍能形成规整球形液滴,而缺失连接区的突变体仅产生无定形絮状沉淀,单独的连接片段甚至比全长蛋白更易形成大量液滴;沉降与浊度定量数据进一步量化差异,缺失连接区蛋白绝大部分留存于上清,几乎不形成可沉淀凝聚物。光诱导 OptoDroplet 系统同步验证该规律,蓝光刺激下除 ΔL 缺失连接突变体外,其余蛋白均可形成细胞内凝聚颗粒,单独连接片段产生的颗粒数量明显更多。最后在大鼠原代海马神经元中重复验证,野生 SNAP25 在神经元胞体与突触区域形成清晰荧光斑点,缺失连接区突变蛋白荧光完全均匀扩散,高分辨成像与己二醇溶解实验均证明点状结构为典型液相凝聚物,完整证明连接区是驱动 SNAP25 发生相分离的唯一核心结构。
3. 棕榈酰化修饰双向调控 SNAP25 相分离与膜锚定
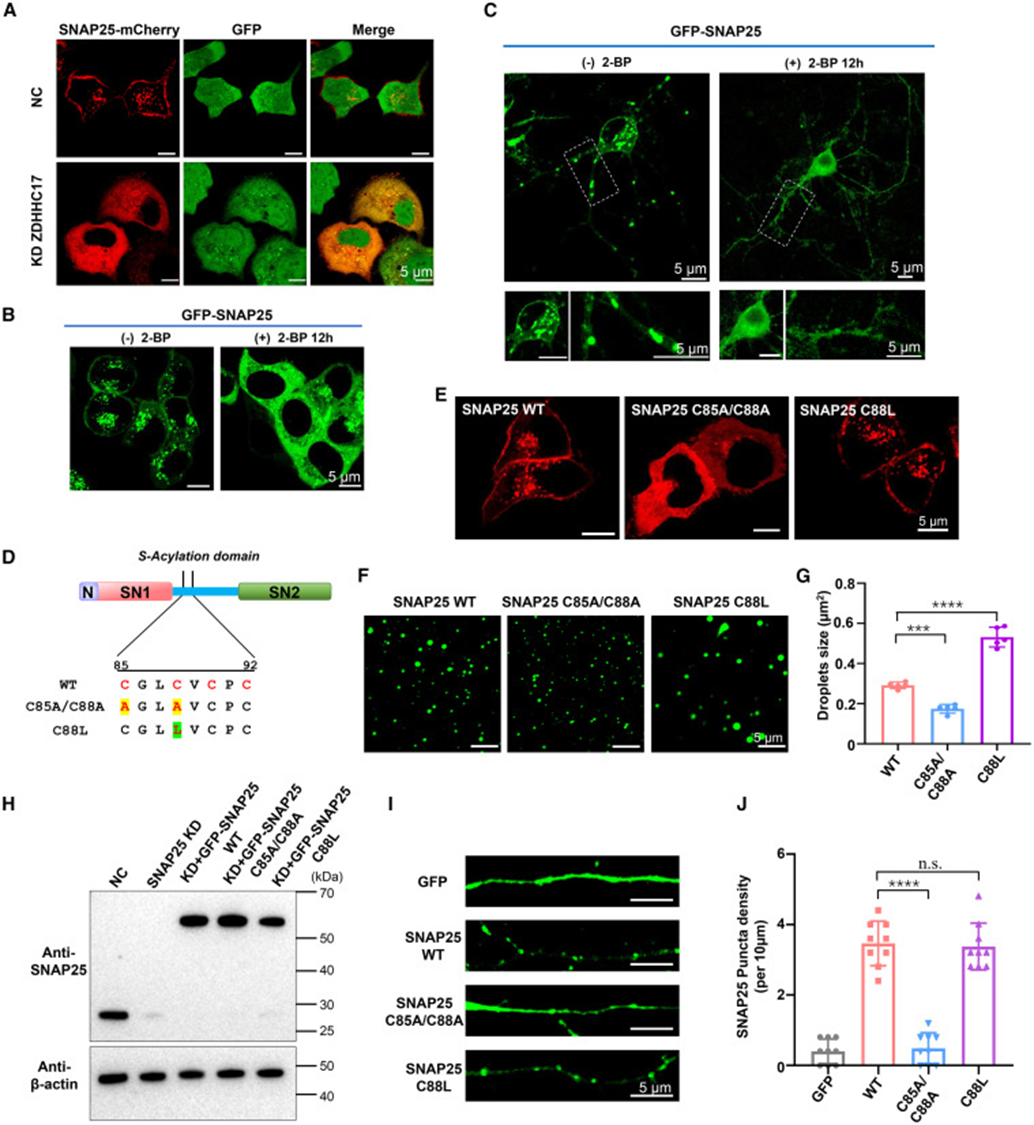
连接区半胱氨酸是棕榈酰转移酶 ZDHHC17 的修饰位点,研究从细胞与体外突变两条路径验证修饰功能。细胞层面敲低 ZDHHC1 或加入棕榈酰化抑制剂 2-BP 后,HeLa 与海马神经元内 SNAP25 点状凝聚物大幅减少,蛋白脱离细胞膜、均匀分布于细胞质,说明棕榈酰化缺失直接破坏凝聚物形成与膜定位。研究者构建两类定点突变:C85A/C88A 双突变消除半胱氨酸修饰位点、C88L 单突变模拟棕榈酰化疏水效应,体外相分离体系中双突变液滴尺寸、数量显著下降,模拟修饰的 C88L 突变液滴更大更密集;平面脂双层 SLB 膜重构实验重现该趋势,只有具备棕榈酰化位点的蛋白才能在膜表面形成高密度蛋白簇,突变蛋白膜聚集能力显著减弱。原代神经元成像同样匹配体外结果,C88L 突变保留大量膜上荧光斑点,而双突变弥散分布,由此证实棕榈酰化通过提升连接区疏水性,一方面促进 SNAP25 液相凝聚,另一方面介导凝聚物锚定在突触细胞膜上。
4. Syntaxin-1 与 SNAP25 共凝聚并增强相分离效率
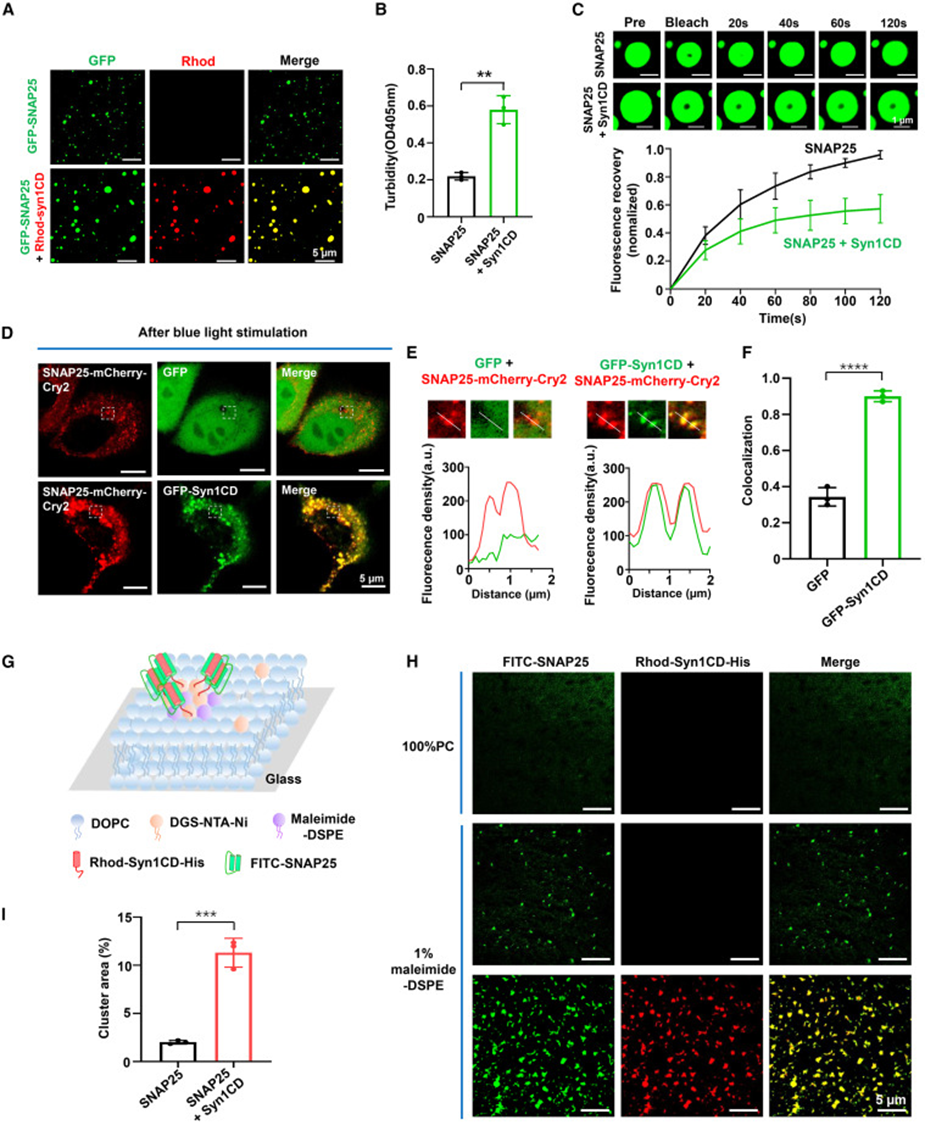
将等量 Syntaxin-1 胞质段与 SNAP25 混合后,双色荧光成像可见两种蛋白完全共定位在同一液滴内,液滴体积显著增大;浊度定量数值同步上升,FRAP 实验显示共凝聚物内部蛋白流动性下降,圆二色谱证明原本无序的 SNAP25 在结合 Syntaxin-1 后转变为 α 螺旋构象,说明二者互作引发蛋白结构折叠。梯度添加 Syntaxin-1 的实验呈现剂量依赖效应,蛋白沉淀中 SNAP25 占比随 Syntaxin-1 浓度持续升高;OptoDroplet 光控共表达体系中,Syntaxin-1 荧光完全渗入 SNAP25 光诱导凝聚颗粒,皮尔森相关系数证明二者高度共定位。人工平面脂双层体系模拟细胞膜环境,膜上固定 Syntaxin-1 后,SNAP25 会大量聚集形成更大共凝聚斑块,从溶液与膜两种环境证明 Syntaxin-1 可与 SNAP25 发生共相分离,同时放大凝聚规模。
5. SNAP25 凝聚物招募全部 SNARE 组分形成三元共凝集体
体外三色荧光标记实验中,Syntaxin-1、VAMP2 两种蛋白均可均匀分布在 SNAP25 液滴内部,沉降实验回收的凝聚沉淀可同时检出三种 SNARE 蛋白,证明凝聚物具备富集两种伴侣蛋白的能力。HeLa 细胞共表达三种荧光标记 SNARE 蛋白时,细胞内生成巨型融合凝聚体,FRAP 结果显示三元共凝集体内部蛋白流动性大幅降低,提示蛋白间广泛相互作用;体外等摩尔混合三种蛋白可直接形成大尺寸共凝聚液滴,配套体外 SNARE 组装实验证实相分离环境下功能性 SNARE 复合体生成量显著提升,明确 SNAP25 液滴作为分子支架,同时捕获两类配套 SNARE 蛋白,为四螺旋融合复合体的组装提供局部高浓度微环境。
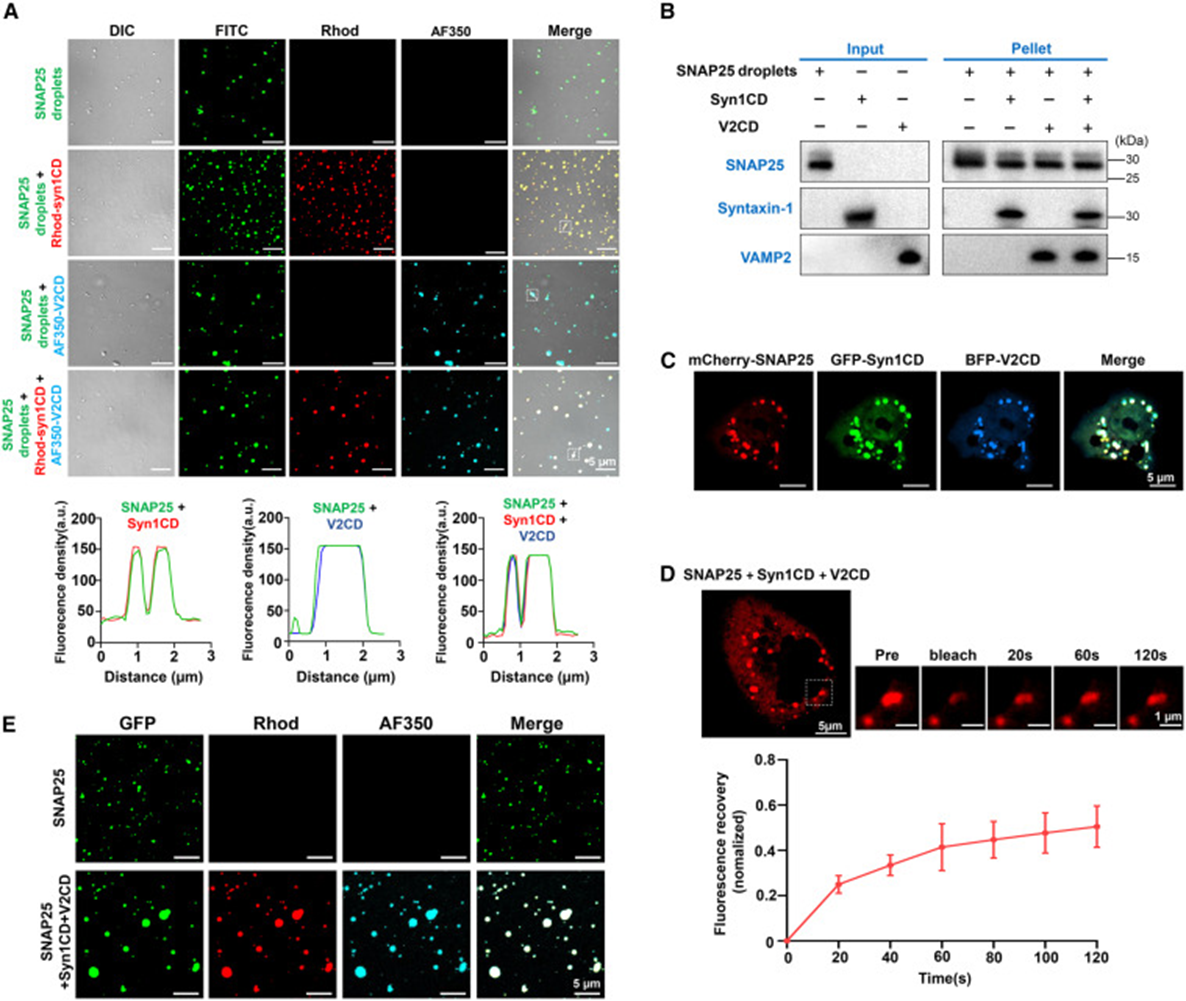
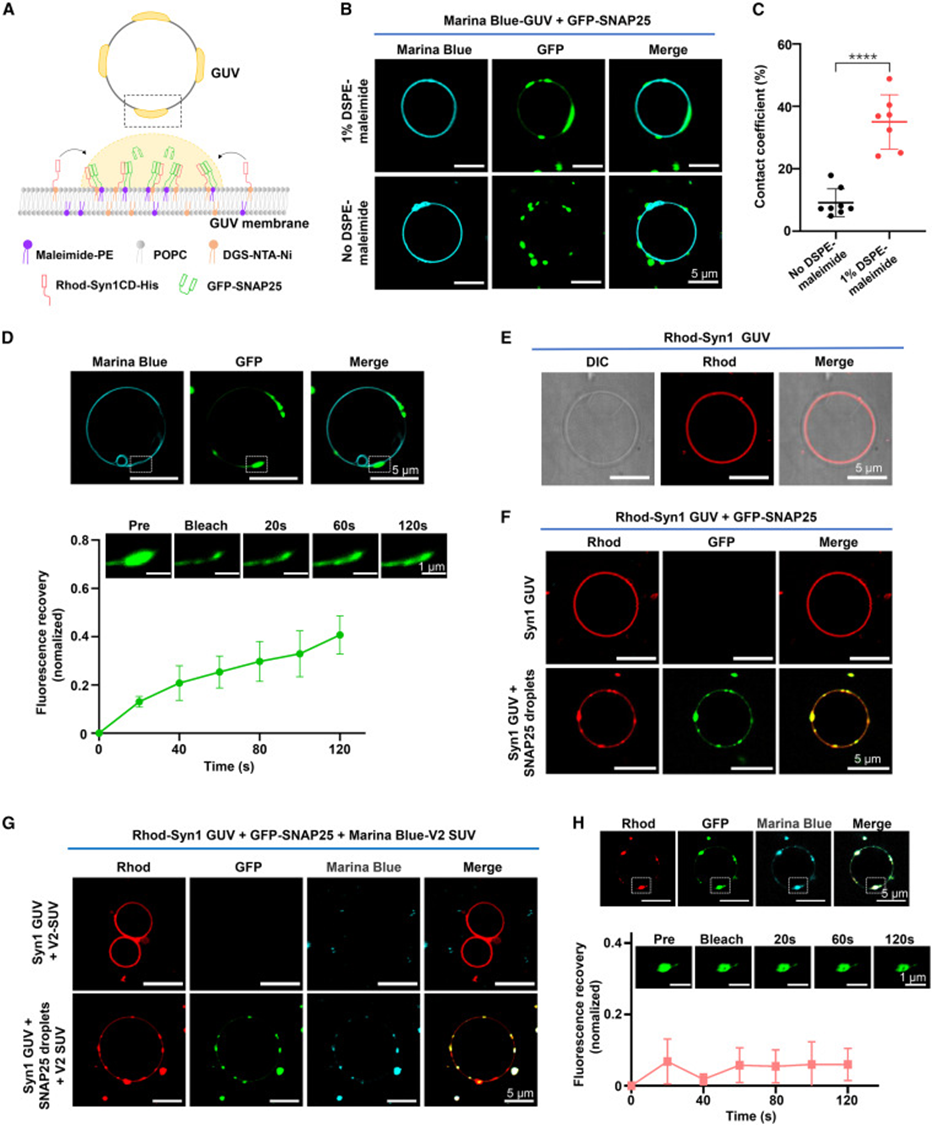
6. SNAP25 凝聚物吸附细胞膜、介导突触囊泡锚定
研究采用 GUV 巨型囊泡模拟突触前细胞膜,带马来酰亚胺脂质的囊泡可结合 SNAP25,荧光成像清晰观察到 SNAP 凝聚物贴附、浸润囊泡表面,若移除马来酰亚胺膜结合位点,液滴无法吸附膜结构。当 GUV 膜预先锚定 Syntaxin-1 时,加入 SNAP25 会在膜表面形成大片共凝聚区域,再加入标记 VAMP 小型囊泡 SUV 后,所有囊泡都会集中富集在膜上 SNARE 共凝聚斑块内,无 SNAP25 时囊泡无法锚定在膜表面。对膜上三元凝聚物开展 FRAP 检测,蛋白流动性明显降低,印证膜、三类 SNARE 多重相互作用稳定囊拴缚状态,完整还原生理过程:膜锚定的 SNAP25 凝聚物搭建融合平台,捕捉携带 VAMP2 的突触囊泡,完成囊泡停靠步骤。
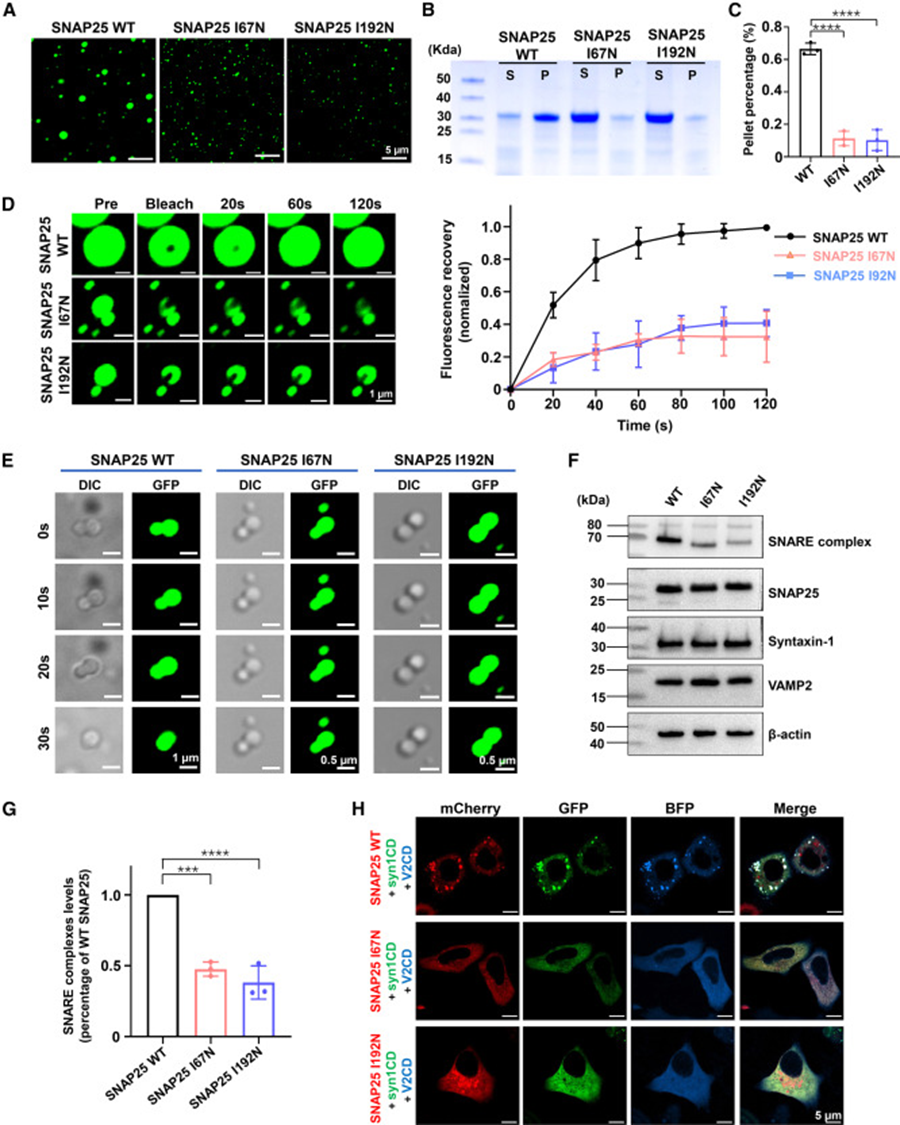
7. DEE 致病突变破坏相分离、抑制 SNARE 复合体组装
选取两种临床 DEE 致病突变 I67N、I192N 开展功能验证,同等体外条件下突变蛋白仅能形成微小、稀少液滴,沉降实验显示进入凝聚沉淀的蛋白量大幅下降;FRAP 曲线证明突变凝聚物内部分子运动受阻,延时液滴融合成像可见突变小液滴几乎无法融合生长,说明突变降低蛋白流动性、阻断液滴融合成熟过程。细胞层面共表达三类 SNARE 蛋白,野生型组可检出大量耐受 SDS 的功能性 SNARE 复合物,两种突变组复合体含量显著下调;三色荧光共转染细胞内,突变 SNAP25 无法与 Syntaxin-1、VAMP2 形成大型三元共凝集体。该组结果建立分子病理通路:致病突变损伤 SNAP25 液液相分离能力,破坏三元 SNARE 共凝聚平台,最终抑制膜融合机器组装,解释 DEE 患者突触释放受损的分子底层机制。
总结
本文围绕突触囊泡融合核心蛋白 SNAP25 展开系统研究,完整揭示棕榈酰化修饰调控的 SNAP25 液液相分离机制,阐明该过程在 SNARE 复合体组装、突触囊泡锚定及神经递质释放中的核心生理功能,并关联发育性癫痫性脑病的发病机理。研究首先通过体外生化与活细胞成像证实 SNAP25 能够发生液液相分离,蛋白两段 SNARE 基序之间的内在无序连接区是实现相分离的必需结构;连接区半胱氨酸棕榈酰化修饰可提升蛋白疏水性,双重促进凝聚体形成与细胞膜锚定,而 Syntaxin-1 可与 SNAP25 共凝聚,进一步放大相分离效应并诱导 SNAP25 构象转变,在膜表面搭建 t-SNARE 富集支架。SNAP25 凝聚体可同步招募 Syntaxin-1 与 VAMP2 形成三元 SNARE 共凝聚体,大幅提升功能性 SNARE 复合体的组装效率,人工膜重构实验证明膜结合的 SNAP25 凝聚体能够浸润膜界面、捕获含 VAMP2 的突触囊泡,解释神经元毫秒级同步递质释放依赖局部 SNARE 蛋白富集的分子基础。研究进一步发现两种 DEE 致病突变 I67N、I192N 会破坏 SNAP25 凝聚体流动性、阻断液滴融合,抑制三元共凝聚体与功能性 SNARE 复合体生成,从分子层面建立相分离缺陷与神经发育疾病的关联。文章客观说明现有局限,尚未解析 Munc18、Munc13 等关键调控蛋白对 SNAP25 凝聚体的调控作用,也缺乏体内动物实验佐证生理功能。整体研究填补了 SNARE 核心蛋白相分离调控突触胞吐的领域空白,提出无膜凝聚体调控快速神经元通讯的全新范式,拓展了突触前分子排布与神经递质释放机制的认知。
EnkiLife恩玑生命产品:
抗体标记试剂盒
Western Blot全流程实验方案
TSA多重荧光试剂盒
细胞荧光染料
稳转细胞系构建服务